



امروز یکشنبه ۲۳ اردیبهشت ۱۴۰۳
دسته بندی سایت
برچسب های مهم
پیوند ها
آمار بازدید سایت

در دهه گذشته، روشن شدهاست که به طور مشابه برای سلولهای هستهدار، گلبولهای قرمز هستهدار (RBCs)مستعد مرگ برنامهریزیشده سلول هستند. به نظر میرسد آپوپتوز اریتروسیتی در پاکسازی فیزیولوژیک گلبولهای قرمز مسن نقش دارد، اما ممکن است در کمخونی منابع علت و معلولی مختلف از جمله دارو درمانی و بیماریهای عفونی دخیل باشد. در مالاریا، کمخونی شدید یک عارضه شایع است که منجر به مرگ کودکان و زنان باردار میشود که در مناطق مالاریا خیز آفریقا زندگی میکنند. پاتوژنز کمخونی مالاریا چند عاملی است و شامل تولید غیر موثر گلبولهای قرمز توسط مغز استخوان و حذف زود هنگام گلبولهای قرمز غیر انگلی شدهاست، پدیدهای که به طور بالقوه با آپوپتوز مرتبط است. در مرور کلی حاضر، ما در مورد شواهد مرتبط با آپوپتوز اریتروسیتی با پاتوژنز کمخونی مالاریایی شدید و همچنین با تنظیم پاک شدن انگل در مالاریا بحث میکنیم.
کلمات کلیدی: مالاریا، کمخونی، آپوپتوز، گلبولهای قرمز خون، فاگوسیتوز
کر و همکارانش اصطلاح آپاپتوز را در سال ۱۹۷۲ برای تعیین یک فرآیند فیزیولوژیک از مرگ برنامهریزیشده سلول که در حفظ نرمال جمعیت سلول بافت و همچنین در توسعه آسیبها مانند سرطان دخیل است، معرفی کردند. از آن زمان به بعد، مطالعات بسیاری با تمرکز بر آپوپتوز منتشر شدهاند. این مطالعات بر آپاپتوز سلولهای هسته ایی متمرکز شدهاند که تغییرات اساسی در هسته، یعنی تراکم کروماتین و تکهتکه شدن DNA، آپاپتوز را به عنوان وابسته به هسته نامگذاری کردهاند. آپوپتوز به عنوان یک فرآیند کلیشهای وابسته به هسته شناخته شد. با این حال، مطالعات پیشرو در مورد سلولهای به طور تجربی انوکلئاسیون شده، نشان داد که ویژگیهای معمول غشا سیتوپلاسمی و آپوپتوز یاختهای، مانند تغییر رنگ غشا، انقباض سلولی، از دست دادن پتانسیل غشایی میتوکندری، و قرار گرفتن فسفاتیدیل سرین میتواند در غیاب یک هسته القا شود، که این احتمال را افزایش میدهد که آپوپتوز میتواند طول عمر سلولهای به طور فیزیولوژیکی انوکلئاسیون شده، یعنی سلولهای قرمز خون (RBC)را نیز تعیین کند.
در واقع، حتی فقدان اندامکها در گلبولهای قرمز، از جمله اندامکهایی که به طور مستقیم در القای آپوپتوزیس دخیل هستند (یعنی میتوکندری و شبکه اندوپلاسمی)، به فعالسازی دستگاه سیتوپلاسمی موجود در سلولی که فرآیند apoptotic را هماهنگ میکند، آسیب نمیرساند. اگرچه مسیرهایی که منجر به apoptosis در RBC میشوند به خوبی شناختهشده نیستند، اما نشانداده شدهاست که آنها با افزایش در Ca۲ + سیتوزولی به دلیل فعالسازی کانالهای کاتیونی غیر انتخابی نفوذپذیر Ca۲ + در غشای سلولی آغاز میشوند، که میتواند توسط انواع زینوبیوتیک ها و مواد درونی تحریک شود.
به نوبه خود، هجوم Ca۲ + منجر به (۱)فعال شدن caspases و calpains میشود، که در جداسازی سلول و تشکیل حباب غشا سلولی مشارکت میکنند؛ (۲)انقباض سلول از طریق خروجی K + از طریق کانال K + حساس به Ca۲ +؛ و (۳)تحریک حرکت تودهای غشا سلولی، منجر به قرار گرفتن فسفاتیدیل سرین (PS)در سطح سلول میشود. این رویداد آخر برای از بین بردن سلولهای apoptotic قبل از فروپاشی سلول و پس از آن آزاد شدن واسطههای پیش التهابی سیتوپلاسمی حیاتی است، زیرا PS به عنوان یک سیگنال "خودخوری" برای سلولهای فاگوسیتی عمل میکند که به شیوهای ضد التهابی عمل میکند.
مشابه آپاپتوز سلولهای هسته دار، آپاپتوز گلبولهای قرمز از طریق پاکسازی فیزیولوژیکی سلولهای پیر در هموستاز بدن شرکت میکند. تخمین زده میشود که ۳۶۰ میلیارد گلبول قرمز بالغ تحت فرآیندهای apoptotic قرار میگیرند و هر روز توسط طحال حذف میشوند، و از عوارض سیستمیک مربوط به همولیز داخل عروقی اجتناب میکنند از سوی دیگر، اعتقاد بر این است که آپاپتوز به کمخونی و اختلالات عروقی مرتبط با شرایط بالینی نیز کمک میکند که در آن میزان بیش از حد گلبولهای قرمز آپاپتوز، از جمله دیابت، نارسایی کلیه، کمخونی سلول داسیشکل، تماس مزمن با سرب و همچنین بیماریهای عفونی مانند سپسیس و مایکوپلاسموز ثبت میشوند.
با توجه به اهمیت پاتوفیزیولوژیک آپوپتوز اریتروسیتی، گروه ما تلاش کردهاست تا این فرآیند اریتروسیتی را در مالاریا مطالعه کند، یک عفونت ناشی از ناقل توسط پروتوزوآ از جنوس پلاسمودیوم که گلبولهای قرمز را آلوده میکند و اختلالات خونی و عروقی قوی را القا میکند. مطالعات ما نشان دادهاند که القای آپوپتوز به گلبولهای قرمز انگلی شده (pRBCs)محدود نمیشود، بلکه همچنین در گلبولهای قرمز غیر انگلی شده (nRBCs)رخ میدهد، و به دخالت هر دو pRBC و nRBC در پاتوژنز مالاریا از طریق احتراق فرآیندهای انتحاری اشاره میکند بنابراین، مرور حاضر شواهدی را پوشش میدهد که دلالت بر آپوپتوز اریتروسیتی در پاتوژنز کمخونی شدید دارد، یک عارضه شایع مالاریا که نشاندهنده یک نگرانی مهم سلامت عمومی است که به شدت با مرگ و میر در کودکان و زنان باردار که در مناطق هیپراندمیک مالاریا در جنوب صحرای آفریقا زندگی میکنند مرتبط است.
کمخونی مالاریایی و حذف گلبولهای قرمز
جای تعجب نیست که کمخونی یکی از شایعترین اختلالات مرتبط با مالاریا است، زیرا انگلهای پلاسموروم یک فرآیند شیزوگونی داخل اریتروسیتی را توسعه میدهند که منجر به لیز گلبولهای قرمز میشود. در کودکان غیر ایمن و بزرگسالان مبتلا به P. falciparum، درجه کمخونی ممکن است با پارازیتمی ارتباط داشته باشد و در بیشتر جوندگان آزمایشگاهی مالاریا، کمخونی حاد نتیجه پارگی درصدهای بالای گلبولهای قرمز خون است. کمخونی حاد میتواند هم در عفونتهای تجربی و هم در عفونتهای انسانی مشاهده شود که پارازیتمی کمی دارند و حتی پس از درمان ضد مالاریا و پاک شدن انگلها، کمخونی میتواند ادامه پیدا کند و یا بدتر شود. با توجه به این حقایق، هم تولید غیر موثر گلبولهای قرمز توسط مغز استخوان و هم حذف زودرس nRBC به عنوان عوامل مهم تعیینکننده پاتوژنز کمخونی مالاریا فرض شدهاند.
میزان حذف RBC در مالاریا در مطالعاتی گزارش شدهاست که تخمین میزنند، برای هر pRBC، تا ۳۲ nRBC از گردش خون حذف میشوند. علاوه بر این، شواهدی وجود دارد که نشان میدهد حذف nRBC به وسیله فعالیت فاگوسیتوز به جای همولیز داخل عروقی میانجی گری میشود ، که سیستم فاگوسیتوز رتیکولواندوتلیال را به عنوان یک مکانیزم مهم پاتوژنز کمخونی مالاریایی برجسته میکند. در واقع، علاوه بر مشارکت مستقیم در کمخونی مالاریایی توسط جذب pRBCs و nRBCs، ماکروفاژها نیز ممکن است با محدود کردن زیست فراهمی سیستمیک آهن بازیابی شده، با فرآیند اریتروپویتیک تداخل کنند، منبع اصلی آن اریتروسیتوزهایی است که در طحال و مغز استخوان رخ میدهد. با این حال، مسیرهای مربوط به پاکسازی گلبولهای قرمز در کمخونی مالاریا، مبهم باقی میمانند و بنابراین، آپوپتوزیس به عنوان یک مکانیزم شناختهشده کاهش سلول شرکتکننده در سیتوپنی چند آسیبی، میتواند موجب حذف گلبولهای قرمز نشده شود.
القای کنندههای آپوپتوز مالاریایی و اریتروسیتی
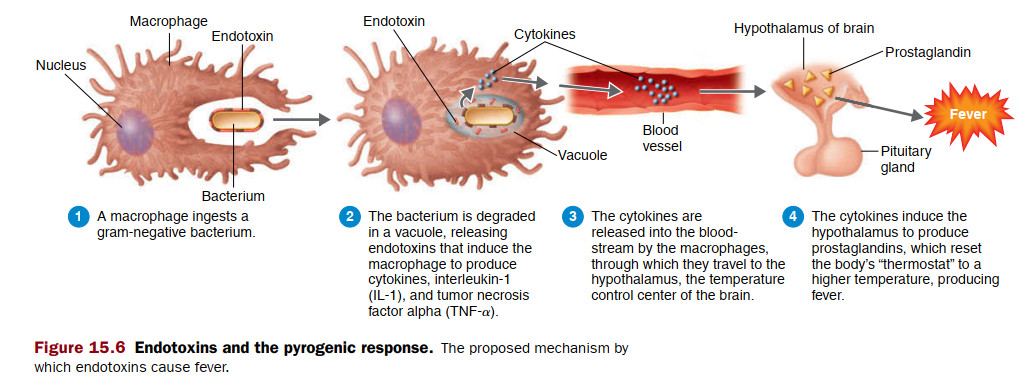
مطالعات انجامشده توسط دوندورپ و همکاران (۱۹۹۹)با بیماران مبتلا به فالسیپاروم P نشان داد که مشابه با pRBCs، که در آن انگلها موجب تغییرات شدید در غشای سلول میزبان میشوند، nRBCs همچنین قابلیت تغییر شکل کاهشیافته مرتبط با شدت کمخونی را ارائه میدهند. این پدیده، که در nRBC از P. falciparum در محیط کشت و موشهای آلوده به P [۲ ] ثبت شدهاست، با جذب آنتیژنهای انگل مالاریا در سطح سلول مرتبط است و جالب توجه است که اخیرا به عنوان مشخصه آپوپتوز اریتروسیتی شناسایی شدهاست و این احتمال را افزایش میدهد که آنتیژنهای انگلهای مالاریا دارای ویژگیهای موافق با apoptotic بر روی گلبولهای قرمز باشند. به طور کلی، میکروب ها میتوانند بر مسیرهای proapoptotic در سلولهای میزبان تاثیر بگذارند. نشانداده شدهاست که باکتریهای گرم منفی و گرم مثبت و همچنین انگلهای تکیاختهای، فرایندهای آپوپتوز سلولهای هستهدار میزبان آلوده و غیر آلوده را از طریق آنتیژنهای پاتوژن دستکاری میکنند که به عنوان مولکولهای تنظیمکننده خزان یاختهای از طریق مسیرهای مختلف عمل میکنند و بر نتیجه بیماری تاثیر میگذارند. تنوع آنتیژنهای تولید شده توسط باکتریهای بیماری زا مانند انتروتوکسین ها، همولیزین، سموم کشنده، اسید لیپوتیکویک و لیپوپلی ساکارید، توانایی افزایش آپوپتوز در انواع مختلفی از سلولهای میزبان از جمله لنفوسیتهای T و B، سلولهای دندریتیک، مونوسیت ها، نوتروفیل ها، سلولهای اندوتلیال، سلولهای اپی تلیال را دارند. به طور مشابه، سموم مالاریایی، گلیکوزیل فسفاتیدیل اینوزیتول و هموزوئن خالص شده از P. falciparum، به ترتیب فعالیت پروپاپتوز در کاردیومیوسیت ها و اریتروبلاست ها را نشان دادند. علاوه بر این، چنین اثر آپاپتوژنیک عوامل انگلی با انکوباسیون سلولهای اندوتلیال عروقی مغز و نوروگلیا با محیط شرطی P falciparum - PRBC ثبت شد، و القای آپاپتوز در سلولهای اندوتلیال ریه میتواند با تماس فیزیکی ناشی از پدیده چسبندگی PRBC رخ دهد. هیچ مطالعهای برای بررسی پتانسیل proapoptotic آنتیژنهای پلاسمودیوال در آپوپتوز اریتروسیتی وجود ندارد.
برچسب های مهم
مطالب تصادفی






/cloudfront-us-east-2.images.arcpublishing.com/reuters/IE22OFLXBRLWZFPBRQMDYEXZVE.jpg)
